
Kmen:
ČLENOVCI (ARTHROPODA)
Název je odvozen z
řec. arthron = kloub, pous, podos = noha. Suchozemští i vodní živočichové, kteří
představují na druhy i jedince nejpočetnější a
evolučně nejúspěšnější živočišný
kmen. Popsán je asi 1 milion druhů, mnohonásobně větší počet druhů je dosud vědě
neznámý.
Hlavní znaky:
článkované končetiny, heteronomní metamerie (nestejnocenně segmentované tělo),
vnější kostra (exoskelet) silně sklerotizovaná, tělní dutina je mixocoel,
otevřená cévní soustava.
Vyjdeme-li z
představy hypotetického "pračlenovce" s homonomní metamerií (stejnocennou
segmentací), pak tyto identické původní články těla různým způsobem splynuly a
vytvořily 3 větší tělní celky - tagmata
(hlava - caput, cephalon, hruď -
thorax, zadeček -
abdomen). Tato
tagmatizace (heteronomní metamerie)
byla jedním z hlavních předpokladů enormní evoluční úspěšnosti členovců.
Coelom
se zakládá embryonálně, později splývá s prvotní tělní dutinou (blastocoelem) a
otevírá se do ventrální cévy, čímž vzniká smíšená tělní dutina -
mixocoel (v anglosaské literatuře
označovaná jako hemocoel). Zbytky coelomu perzistují jen v urogenitálním
systému.
Cévní soustava
je otevřená. Dorzální pulzující
céva (aorta) bočními otvory (ostia)
nasává hemolymfu (krvomízu), žene ji
kraniálně, kde ji vytlačuje do mixocoelu. Bývá někdy označována jako "srdce".
Končetiny jsou
článkované, články jsou spojené
klouby, kterými jsou končetiny také
připojeny k tělu. Tyto klouby jsou však jiného typu než u obratlovců. Původně
každý článek těla nesl 1 pár končetin, z nichž některé v průběhu evoluce
nabyly jiné funkce. To je nejlépe patrné na hlavových článcích (viz tab. 1).
Končetiny původního typu byly dvojvětevné, tak jak se zachovaly pouze u korýšů.
U ostatních recentních podkmenů vnější větev vymizela a končetina se skládá z
kyčle (coxa), příkyčlí (trochanter),
stehna (femur), holeně (tibia) a
chodidla (tarsus) zakončeného obvykle dvěma drápky, někdy přísavkou
(zákožky).
Tab.
Přehled modifikovaných končetin na hlavových článcích 3 podkmenů členovců.
|
klepítkatci
(Chelicerata) |
korýši
(Crustacea) |
šestinozí
(Hexapoda) |
|
chelicery |
tykadla 1.
řádu - antenuly |
tykadla -
anteny |
|
pedipalpy |
tykadla 2.
řádu - anteny |
kusadla -
mandibuly |
|
kráčivé
končetiny (4 p.) |
kusadla -
mandibuly |
čelisti -
maxily |
|
|
čelisti -
maxily 1. řádu |
spodní pysk
- labium |
|
|
čelisti -
maxily 2. řádu |
kráčivé
končetiny (3 p.) |
|
|
další
končetiny |
|
Tělo je kryto
kutikulou, která nasedá na bazální
membránu. Bázi kutikuly tvoří jednovrstevný cylindrický nebo kubický epitel,
tzv. hypodermis, na ni nasedá poměrně silná endodermis, dále ektodermis a
konečně epidermis. Kutikula je složená převážně z bílkoviny
sklerotinu, jehož součástí je
chitin. U různých skupin pak dochází
ke sklerotizaci a inkrustacím ukládáním CaCO3. Vnější kostra brání
kontinuálnímu růstu, a proto dochází k opakovaným svlékáním -
ekdyzím, jimiž se členovci nebo
jejich larvy zbavují svrchních vrstev kutikuly, někdy i metabolitů. K ekdyzi
dochází mezi endodermis a ektodermis. Svlečka se nazývá
exuvie.
Kutikula
na povrchu vytváří pevnou vnější kostru -
exoskelet a její výběžky zasahují do
nitra tagmat a vytvářejí tam částečnou vnitřní kostru - endoskelet. Vnější
kostra se zanořuje i do některých tělních otvorů (stigmata) jako výstelka
trachejí (intima neboli taenidium), která je také svlékána. Každý článek je
svrchu kryt hřbetní destičkou - tergum
resp. tergit a břišní destičkou
- sternum resp.
sternit. Obě destičky jsou spojeny měkkými a pružnými blankami -
pleurami, stejně jako klouby
končetin. Při růstu těla se pleura může do jisté míry roztahovat. S vnější
kostrou souvisí limitovaná maximální velikost členovců – u recentních forem
nanejvýš desítky cm (s končetinami i více, např. velekrab japonský – 3,5 m). Při
větší velikosti těla by byl exoskelet příliš těžký a nefungovala by výměna plynů
vzdušnicemi - proto největších velikostí dosahují vodní členovci, kteří jsou
nadnášeni a nedýchají trachejemi.
Obr.
Schéma tělního článku členovce. (Převzato z Obenbergera, 1952)
Svalovina
členovců je příčně pruhovaná, kromě
svaloviny trávicí soustavy, která je hladká. Svaly pohybového aparátu se upínají
na vnější kostru zevnitř.
Trávicí soustava
je úplná a je tvořena přední (stomodeum),
střední (mezodeum) a zadní částí (proktodeum).
Přední a zadní část jsou původu ektodermálního a jsou vystlány kutikulou,
zatímco nejdelší střední úsek je entodermálního původu, vystlaný jednovrstevným
cylindrickým epitelem s trávicí schopností. Jednotlivé úseky jsou pojmenovány
analogicky s obratlovci. Stomodeum se skládá z dutiny ústní, hltanu, jícnu,
event. volete a žvýkacího žaludku. Slouží k mechanickému zpracování potravy. Je
zakončeno svěračem (valvula cardiaca). Mesodeum zahrnuje žlaznatý žaludek,
kraniální úsek střeva, který je oddělen od proktodea svěračem (valvula
pylorica). Proktodeum představuje kaudální část střeva a konečník.
Přídatnou žlázou
trávicího traktu je slinivkojaterní žláza -
hepatopankreas.
Vylučovací ústrojí
představují různě modifikovaná párová
metanefridia, jako antenální žlázy
ústící při bázi tykadel, koxální žlázy
při bázi končetin nebo maxilární žlázy
u čelistí. U hmyzu a některých pavoukovců jsou
malpighické trubice (podle italského
přírodopisce M. Malpighiho, 1628 – 1694).
Dýchací aparát
je utvářen u jednotlivých podkmenů odlišně. Suchozemští členovci dýchají
vzdušnicemi (trachejemi),
plicními vaky nebo obojím. Korýši dýchají
žábrami, u některých vodních larev
hmyzu se vytvářejí tracheální žábry,
což jsou tracheje vychlípené do vodního prostředí. Nejmenší členovci s jemnou
kutikulou (např. někteří roztoči) dýchají
celým povrchem těla.
Nervová soustava
je žebříčková (viz kroužkovci), je
zde však mnohem výraznější cefalizace.
Tak jak docházelo ke splývání článků hlavy, tak také došlo ke splynutí hlavových
ganglií, která vytvořila nadjícnovou resp. nadhltanovou (supraezofageální) a
podjícnovou (subezofageální) zauzlinu, objímající spojkami jícen. Z podjícnové
zauzliny po břišní straně kaudálním
směrem pokračuje „žebříček“ resp. břišní
nervová páska (rozdílná poloha nervového systému oproti strunatcům). Na
nadjícnovou uzlinu se napojuje systém
endokrinních žláz řídících především ontogenetický vývoj. Je uspořádán
funkčně hierarchicky obdobně jako u obratlovců osa hypotalamus - hypofýza -
nadledviny aj. žlázy (podrobněji viz u hmyzu).
Nervový systém je na
periferii zakončen složitými smyslovými
ústrojími. Základem smyslových ústrojí (zejm. mechanoreceptorů) je
sensila (sensillum), receptorová
buňka s jemnou brvou (seta),
pronikající kutikulou na povrch těla a přenášející změny např. pohybu do nervové
buňky. Sety mohou sloužit i k detekci zvuku. Změny tlaku a vibrace vzduchu a
vody jsou vnímány chordotonálními orgány,
což jsou sensily registrující změnu napětí kutikulárních strun. Zvuky vnímají
jen někteří členovci (blíže viz u hmyzu).
Oči
členovců jsou v zásadě dvojího typu. Buď jsou to
oči
jednoduché (ocelli), které se
vyskytují i u larev a u dospělců jako očka temenní nebo jsou to nejčastěji
oči
složené (oculi compositi), tvořené
velkým počtem jednoduchých ommatidií.
Členovci, zvláště hmyz, mají schopnost vnímat i polarizované světlo nebo UV
paprsky, což má zvláštní význam pro orientaci hmyzu při oblačné obloze.
Univerzálním atraktantem pro krevsající členovce (klíšťata, komáři aj.) je CO2
vydechovaný obratlovcem, mohou to však být i těkavé mastné kyseliny z jeho potu.
Pohlavní orgány
jsou párové, trubicovité stavby. Často je vytvořena spermatéka (receptaculum
seminis), která umožňuje uchovávat životaschopné spermie i po několik let
(včela).
Rozmnožování
členovců je pohlavní, převážná většina jich jsou
gonochoristé (výjimkou jsou korýši
svijonožci). Oplození je obvykle vnitřní.
Existuje zde oviparie, ovoviviparie i viviparie. U členovců není vzácná
partenogeneze,
neotenie resp.
pedomorfóza (rozmnožování roztočů v
larválním stadiu). Jiným typem rozmnožování je
heterogonie (střídání partenogeneze
s rozmnožováním pomocí oplozených vajíček např. u perlooček nebo mšic).
Příkladem nepohlavního rozmnožování je velmi vzácná
polyembryonie, vyskytující se jen u
některých parazitických blanokřídlých.
Vajíčka
členovců jsou
mozaiková neboli
determinační. To umožňuje také vznik tzv.
gynandromorfů, jedinců, jejichž každá strana náleží jinému pohlaví. Jelikož
jsou vajíčka mezo- nebo
polylecitální, dochází k povrchovému
(superficiálnímu) rýhování. Některé velmi primitivní formy mají vajíčka
oligolecitální a pak dochází k
totálnímu rýhování.
Vývoj
členovců je buď přímý (např. pavouci, raci) nebo nepřímý, tj. přes
larvu.
Členovci se
klasifikují do několika podkmenů, které jsou morfologicky definovány především
uspořádáním přeměněných končetin na hlavových článcích. Dřívější podkmen
vzdušnicovci (Tracheata) jakožto nepřirozený taxon se již většinou neuvádí.
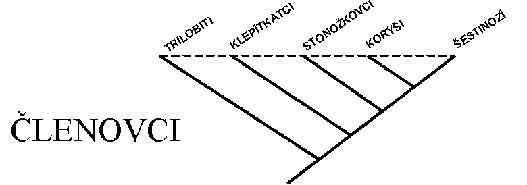
Stonožkovci jako jeho součást jsou podle molekulárně fylogenetických studií
blíže příbuzní klepítkatcům než šestinohým resp. hmyzu, a ti jsou opět
příbuznější korýšům než stonožkovcům.
Členovci jsou díky
obrovskému počtu jedinců důležitou součástí potravních řetězců na souši i v
mořích. Např. mravenci tvoří téměř 30 % živočišné biomasy v Amazonii a mořský
korýš krunýřovka krillová tvoří 2x větší biomasu než celé lidstvo. Další význam
členovců je podrobněji zmíněn u hmyzu.
Kromě fosilních
trilobitů se členovci dělí na čtyři recentní podkmeny:
Podkmen: Klepítkatci
(Chelicerata)
Podkmen: Stonožkovci
(Myriapoda)
Podkmen: Trilobiti
(Trilobitomorpha)
Vymřelí prvohorní
mořští živočichové, jejichž tělo bylo rozděleno podélně i příčně na tři části
(lat. lobus = lalok). Rozeklané končetiny byly na zevní větvi opatřeny žábrami.
Velikost byla několik cm, výjimečně až 70 cm. Jejich postavení v systému
členovců je nejasné (vztah k ostrorepům). Typičtí živočichové pro barrandién
(geologický útvar v Čechách, podle paleontologa francouzského původu J.
Barranda, 1799–1883) .
Tyto výukové materiály byly spolufinancovány Evropským sociálním fondem a státním rozpočtem ČR.