
Individuální vývoj
(ontogeneze) živočichů začíná
zpravidla oplozením vajíčka a končí smrtí živočicha. Probíhá ve dvou obdobích –
embryonálním a postembryonálním (mezi něž bývá u savců včleněno ještě tzv.
období fetální).
Během embryogeneze je jedinec chráněn vaječnými obaly či tělem matky a je tedy relativně nezávislý na vnějším prostředí. Oplozené, případně aktivované apomiktické, vajíčko se v období rýhování mitoticky dělí na množství blastomer. Regulační vajíčka, vajíčka s funkčně nerozlišenou cytoplazmou, dávají vznik funkčně nerozlišeným blastomerám (je tu možná polyembryonie). Determinační (mozaiková) vajíčka mají cytoplazmu rozlišenou a každá ze vznikajících blastomer je již determinována pro vznik určité části těla budoucího jedince. Vajíčka se mohou lišit množstvím a rozložením žloutku. Vajíčka alecitální jsou zcela bez žloutku (někteří ploštěnci), oligolecitální mají žloutku málo (živorodí savci), mezolecitální více (obojživelníci) a velmi bohatá na žloutek jsou vajíčka polylecitální (plazi, ptáci). Ve vajíčkách izolecitálních je žloutek rozložen v cytoplazmě rovnoměrně, také rýhování je rovnoměrné (ekvální) a vzniklé blastomery jsou stejně velké (živorodí savci). Naopak vajíčka anizolecitální mají žloutek soustředěn buď k vegetativnímu pólu (vajíčka telolecitální, např u ptáků) nebo žloutek vyplňuje střední část vajíčka a cytoplazma je na povrchu (vajíčka centrolecitální, např. u hmyzu). Rýhování je pak nerovnoměrné (inekvální), tj. blastomery na animálním pólu jsou menší a na vegetativním větší (obojživelníci). Při úplném (totálním) rýhování se rýhuje celé vajíčko (vajíčka holoblastická). Pouze polylecitální anizolecitální vajíčka se rýhují jen částečně (vajíčka meroblastická) tzn., že jejich střed (centrolecitální vajíčka hmyzu – rýhování povrchové, superficiální) nebo vegetativní pól zůstává nerozrýhován (telolecitální vajíčka plazů a ptáků – rýhování terčovité, diskoidální, rýhuje se jen animální pól).
Obr. Gastrulace
ptačího embrya. Meroblastickým rýhováním vzniká blastoderm (A, B), který se
rozštěpí na epiblast a hypoblast (C). Centrem organogenní aktivity je tzv.
primitivní proužek vyznačující předozadní osu zárodku. Emigrující buňky
epiblastu dávají vznik ento- a mezodermu (červeně) (D). Postupně se formují
nervová trubice, střevo, chorda a somity (červeně) (E) (JK podle Millera a Harleye, 1996).
Z hlediska
symetrie rozeznáváme rýhování radiální,
kdy rýhy probíhají v poledníkových a rovnoběžkových rovinách (druhoústí) a
spirální, kdy jsou blastomery
v řadách nad sebou posunuty, jako když klademe cihly při stavbě zdi (prvoústí).
U savců však dochází k tzv. rotačnímu
rýhování, které připomíná spirální rýhování prvoústých.
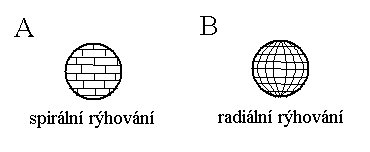
Obr.
Schématické srovnání rýhování vajíčka u
prvoústých (A) a druhoústých živočichů (B) (Obr. OS).
V průběhu
blastogeneze dále prochází zárodek
stadiem moruly,
blastuly a
gastruly.
Morula (podle podobnosti s plodem moruše) je seskupením blastomer.
Blastula vzniká z moruly zmenšením a středovým rozestupem blastomer. Její
stěna je tvořena jednou vrstvou epitelově uspořádaných buněk (blastodermem),
který uzavírá prvotní tělní dutinu -
blastocoel.
Morfogenezí, tzv. morfogenetickými
pohyby (invaginace, imigrace) se jednovrstevná blastula mění ve dvouvrstevnou
gastrulu, vytváří se ektoderm a
endoderm (entoderm) (vnější a
vnitřní zárodečný list). Vnitřní prostor gastruly (prvostřevo, archenteron) je
spojen s vnějším prostředím otvorem tzv. prvoústy (blastoporem).
U většiny živočichů se vytváří také třetí zárodečný list (parenchymatický
mezenchym nebo pravý
mezoderm). Vznikem zárodečných listů dochází také ke změnám tělní dutiny. U
žahavců nahrazuje blastocoel řídká rosolovitá buněčná síť mezi ekto- a
entodermem, tzv. mezoglea. U
bilaterií se tradičně uváděly 3 hlavní
typy tělních dutin - schizocoel, pseudocoel a coelom (célom). U ploštěnců je
tělní dutina omezena na soustavu štěrbinek v parenchymatickém mezenchymu
vyplněných tělní tekutinou, což se tradičně označovalo jako
schizocoel. U hlístic se štěrbiny
rozestupují a vytvářejí prostornou dutinu mezi ekto- a entodermem vyplněnou
tekutinou, tzv. pseudocoel.
U kroužkovců nebo strunatců se druhotná tělní dutina (coelom)
vytváří uvnitř mezodermálních
coelomových váčků. Ty vyplňují prostor mezi ekto- a entodermem, a tak
výrazně potlačují prvotní tělní dutinu. Tím dochází i k uzavření tělní tekutiny
do krevního oběhu. Vznik coelomu je tak spjat s existencí pravého mezodermu. U
členovců se coelomové váčky druhotně rozpadají, a tak se spojují zbytky prvotní
tělní dutiny s dutinou druhotnou v tzv.
mixocoel.
U většiny
živočišných kmenů plní prvoústa gastruly v pozdějších fázích ontogeneze funkci
ústního otvoru a u skupin s úplnou trávicí soustavou vzniká na opačném konci
zárodku otvor řitní (tzv. prvoústí –
Protostomia). U strunatců a několika dalších kmenů prvoústa zanikají, ústní
otvor se druhotně vytváří na protějším pólu zárodku a řitní otvor vzniká
v oblasti prvoúst (tzv. druhoústí –
Deuterostomia).
U strunatců se během
embryonálního vývoje coelomové váčky dělí v párové
somity umístěné dorzolaterálně
a nepárový splanchnotom na ventrální
straně. Struktury vznikající ze segmentovaných somitů jsou tedy primárně
segmentované a nazývají se somatické – např. somatické nervy inervující ústroje
somatického původu. Struktury odvozené od nesegmentovaného splanchnotomu se
nazývají splanchnické nebo viscerální – např. viscerální nervy inervující
ústroje viscerálního původu. Z vnější části somitu zvané
dermatom vzniká škára (dermis),
z vnitřní části zvané myotom vzniká
svalovina a z vyčleněných skupin buněk zvaných
sklerotom vzniká skelet.
Z dorzálních partií splanchnotomu zvaných
nefrotom a
gonotom vznikají základy
urogenitální soustavy. Epitel kryjící vnitřní orgány se nazývá
splanchnopleura a stěnu tělní dutiny
vystýlá somatopleura. Během evoluce
strunatců se postupně vytrácí segmentace coelomových váčků a orgánů.
Diferenciací buněk
zárodečných listů vznikají základy orgánů, ze kterých se při
organogenezi vyvíjí orgány. Původ
nejdůležitějších orgánů: 1) orgány
ektodermálního původu – pokožka s kožními žlázami a její deriváty, smyslové
orgány, nervová soustava, malpighické trubice a tracheje vzdušnicovců,
protonefridie, plícní vaky pavoukovců, přední a zadní část trávicí trubice;
2) orgány mezodermálního původu
– svalová soustava, pojiva, metanefridie, ledviny obratlovců, pohlavní orgány,
cévní soustava; 3) orgány entodermálního
původu – trávicí trubice (kromě přední a zadní části) a její přídatné žlázy,
struna hřbetní, dýchací orgány (plíce, plynové měchýře) obratlovců. Po ukončení
organogeneze se embryo savců označuje jako plod (foetus,
fetus). Embryonální, resp. fetální období končí vylíhnutím či porodem (partus).
a

b

c

d

e

f

g

h

i

j

Obr. Jednotlivé fáze porodu siky vietnamského.
V porodních cestách se objevují spárky a čenich plodu (a), matka uléhá a vstává
(b, c), mládě je vypuzeno (d) a padá na zem (e), je dosucha olizováno matkou (f,
g), objevují se plodové blány s plodovou vodou (h), matka požírá placentu (i) a
mládě se zanedlouho již pokouší vstát (j) (Foto IL).
V závislosti na množství žloutku ve vajíčkách, případně na možnosti výživy zárodku z těla matky, opouštějí juvenilní jedinci vaječné (plodové) obaly v různém stadiu ontogeneze. Během evoluce se u živočichů vyvinuly dvě základní strategie zajišťující úspěch jejich reprodukce. První je založena na produkci velkého počtu vajíček s malým množstvím žloutku a bez složitých obalů. Embryonální vývoj je krátký a vajíčko opouští velmi primitivní, tzv. primární, palingenetická larva, která je obvykle velmi odlišná od dospělců a často jde jen o modifikovanou gastrulu (např. planula žahavců, Müllerova larva některých ploštěnek, trochofora kroužkovců a primitivních měkkýšů, pilidium pásnic, veliger mořských plžů a mlžů, nauplius korýšů, larvy ostnokožců). Tyto larvy mohou být lecitotrofní (z řec. lekithos = žloutek), tj. spotřebovávají žloutek a nepřijímají potravu, nebo planktotrofní (živí se planktonem). Druhou strategií je produkce menšího počtu vajíček s velkým množstvím žloutku a s ochrannými obaly. Vývoj je delší, zárodek se líhne dokonaleji vyvinut a teoreticky s větší nadějí na přežití. V tomto případě opouští vajíčko buď vývojově dokonalejší, tzv. sekundární, cenogenetická larva, která má již složitou stavbu a v mnoha rysech připomíná dospělce (larvy hlístic, zoëa korýšů, larvy hmyzu, minoha mihulí, plůdek ryb, pulci obojživelníků aj.), nebo se z vajíčka líhne mládě. V případech, kdy vývoj jedince probíhá přes jakýkoli typ larvy, hovoříme o vývoji nepřímém. V parazitologii je však často jako nepřímý vývoj chápán vývoj s mezihostitelem, jako přímý vývoj bez mezihostitele. U hmyzu rozlišujeme ještě vývoj nepřímý s proměnou nedokonalou (hemimetabolie – larvy se podobají dospělci – imagu; kobylky, ploštice, všenky, aj.) a s proměnou dokonalou (holometabolie – larvy se morfologicky i ekologicky liší od imága, které vzniká metamorfózou přes stadium kukly; tzv. kampepoidní larvy jepic a vážek, ponravy brouků, housenky motýlů, housenice pilatek, strusky dvoukřídlých, aj.).

Obr. Hemimetabolie - nymfy cikády (Foto JS).

Obr. Holometabolie. Larvy motýlů (na obr.
housenka lišaje kyprejového) procházejí dokonalou proměnou, jejíž evoluční
výhodou oproti proměně nedokonalé je zřejmě odstranění potravní konkurence mezi
larvou a dospělcem (imagem) (Foto OS).
Svou stavbou
stojí larvy holometabolního hmyzu někde mezi primárním a sekundárním typem
larev, obdobně jako např. glochidium
mlžů či onkosféra tasemnic. Pokud se
líhne nebo rodí mládě, které se od dospělce liší jen velikostí, jde o
vývoj přímý. Nejdokonalejším
případem je vývoj uvnitř mateřského organismu, který zárodek chrání a mnohdy i
vyživuje (zárodečné obaly – amnion,
seróza neboli chorion, allantois – pouze u obratlovců). Pravděpodobnost
přežívání jedince je často zvýšena rodičovskou péčí o potomky. Nejčastěji se
noví jedinci líhnou z vajíček uložených v prostředí mimo tělo matky (vejcorodost,
oviparie). Někdy však zůstávají vajíčka v pohlavních vývodech matky a její
tělo opouštějí již larvy nebo juvenilní jedinci (vejcoživorodost,
ovoviviparie – některý hmyz, ryby, plazi, např. zmije obecná). Zvláštní typy
nepravé ovoviviparie se vyskytují u některých ryb, např. tlamovci zadržují
vajíčka až do dokončení vývoje zárodku ne ve vývodech pohlavních cest, ale
v ústech; dále u některých žab, např. u pipy americké se vyvíjejí vajíčka
v komůrkách zduřené hřbetní kůže samice. Samice australské žáby
Rheobatrachus oplozená vajíčka
spolknou. Pulci, kteří se z nich vylíhnou, absolvují svůj vývoj v žaludku, který
po dobu růstu pulců ztrácí svou trávicí funkci. Výjimečně dochází k larvorodosti
(larviparii), kdy larvy prodělávají
část vývoje v těle samice (někteří roztoči a dvoukřídlí, mlok skvrnitý). Jsou-li
vajíčka nebo zárodky v průběhu vývoje vyživovány alespoň částečně z těla matky
prostřednictvím speciálních tkání (různé typy placenty), jde o živorodost (viviparii
– známá je u obratlovců, nejdokonalejší je u placentálních savců, ale existuje i
u některých bezobratlých – mšice, švábi. U některých dvoukřídlých dokonce
vznikají v těle matky larvy, které se živí jejími tkáněmi a poté přezimují ve
zbylé zevní kutikule.
Postembryonální
období
lze rozdělit na tři etapy:
prereproduktivní (juvenilní),
reproduktivní (fáze dospělosti) a postreproduktivní
nebo senektivní (fáze stárnutí).
Délka uvedených období je u jednotlivých skupin živočichů různá. U bezobratlých
obvykle výrazně převažuje svojí délkou období prereproduktivní, u většiny
obratlovců je nejdelší částí života období reproduktivní a jen v některých
případech zahrnuje delší dobu i období postreproduktivní (velcí ptáci a savci).
Tělesný růst obvykle ustává s přechodem do reproduktivního období (savci, brouci
aj.), ale někdy pokračuje až do smrti (ryby, plazi).
Tyto výukové materiály byly spolufinancovány Evropským sociálním fondem a státním rozpočtem ČR.