Podle nových
poznatků mají živočichové (Metazoa) monofyletický původ a vyvinuli se
z jednobuněčných eukaryotních organismů, konkrétně ze společného předka s
trubénkami (Choanoflagellata). Jedním z důvodů vzniku mnohobuněčnosti byl fakt,
že nebylo možné zvětšovat velikost buněk nad určitou mez, danou prostými
fyzikálními zákonitostmi. Jedná se o
vztah mezi povrchem a objemem koule při zvětšujícím se poloměru. Difúze
plynů a rozpuštěných látek je efektivní jen při relativně velkém povrchu; s
růstem poloměru relativní velikost povrchu klesá a výměna se stává nefunkční.
Proto např. celým povrchem těla dýchají především nejmenší živočichové a proto
bylo nutné jít cestou zmnožení počtu buněk při zachování jejich malé velikosti.
Větší velikost přináší řadu výhod a ve vývojových liniích živočichů lze
vysledovat tendenci ke zvětšování velikosti. Možnou cestu od jednobuněčnosti
k mnohobuněčným živočichům se snažilo vysvětlit několik spekulativních hypotéz,
které formulovali významní biologové před více než 100 lety (E. Haeckel, I.
Mečnikov, O. Bütschli aj.). Uchylovali se přitom k analogiím s existujícími
organismy a embryonálním vývojem. E. Haeckel je autorem tzv.
biogenetického zákona, který praví,
že ontogeneze je zkráceným opakováním fylogeneze. Toto pravidlo je dnes často
pokládáno za neplatné, i když najdeme případy, na které je lze aplikovat. Ze
zmíněných hypotéz zaslouží zmínku tzv. koloniální hypotéza předpokládající,
že na počátku vývoje živočichů byly kolonie jednobuněčných bičíkatých "prvoků"
nebo tzv. plakulární hypotéza inspirovaná vločkovci (kmen Placozoa). První
živočichové byli mořští, pravděpodobně bentičtí, s povrchovou vrstvou obrvených
(monociliátních) buněk a vnitřními kmenovými buňkami bez bičíků. Postrádali
ještě zřejmě symetrii. Z recentních živočichů se této představě nejvíce blíží
právě vločkovci. Postupně pak docházelo k diferenciaci buněk do tkání, orgánů a
specializovaných soustav.
Ačkoli se v poslední
době díky molekulárně biologickým metodám začínají vyjasňovat skutečné
fylogenetické vztahy mezi jednotlivými skupinami živočichů, mnohé aspekty
fylogeneze této říše zůstávají nadále zahaleny tajemstvím a jsou předmětem
spekulací. Jisté je, že si vývojový „strom života“ nemůžeme představovat jako
hlavní kmen s postranními větvemi, který směřuje vzhůru k obratlovcům, savcům a
posléze k člověku jakožto svému cíli a vrcholu, jak to bývá někdy mylně chápáno.
Výstižnějším a správnějším schematickým vyjádřením evoluce jsou do různých stran
se rozbíhající a větvící vývojové linie, představující různé alternativní cesty.
Pokud jako kritérium evoluční úspěšnosti bereme početnost populace a její
vzrůstající tendenci, pak je člověk ve své velikostní kategorii skutečně
nejúspěšnější; nemůže se ovšem měřit např. s hmyzem. Pokud za evoluční úspěšnost
vezmeme délku přežívání na této planetě, pak se jako jedna z nejúspěšnějších
skupin jeví např. ostnokožci, kteří přežili 540 milionů let v dodnes velkých
počtech, a to navzdory opakovaným katastrofám, jež přivodily zánik většiny
jejich současníků. Savci oproti nim obývají Zemi relativně krátkou dobu (přes
200 mil. let) a moderní lidé (Homo
sapiens) existují teprve 200 tis. let.
Jedním ze základních mechanismů evoluce je adaptivní radiace, dříve častěji označovaná jako divergence (rozbíhavost) vývojových linií. Název radiace (doslova vyzařování) naznačuje, že jde o paprsčité větvení původně jediné vývojových linie (společný předek). Ve skutečnosti jde ovšem o opakované dichotomické větvení. Přívlastek adaptivní říká, že se tak děje v procesu adaptace, tj. přizpůsobování se rozdílným podmínkám prostředí. Dochází k obsazování volných ekologických nik a rozrůznění původně vzájemně si podobných forem, které na druhé straně mohou připomínat jim nepříbuzné živočichy žijící v obdobných podmínkách. Příkladem je rozrůznění australských vačnatců ze společného předka připomínajícího krysu v tak rozdílné živočichy jako je vakokrt, vakoveverka, vakovlk nebo klokan. Tato jejich podobnost s placentálními savci z jiných kontinentů je příkladem konvergence (sbíhavost znaků). Jde o vnější podobnost vývojově nepříbuzných forem způsobenou adaptací na obdobné podmínky prostředí. Klasickým příkladem tvarové konvergence je podobnost kytovců s rybami nebo např. podobnost mravenců a termitů.
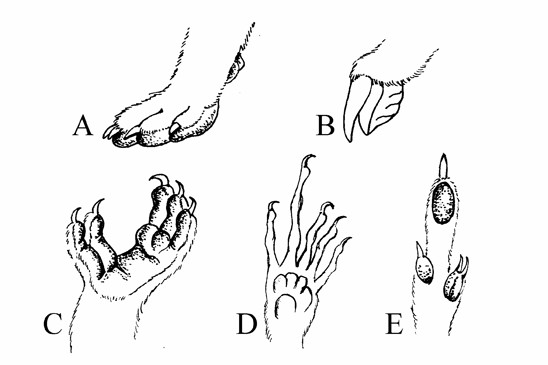
Obr. Konvergence (a současně adaptivní radiace) na příkladu končetin vačnatců: A - noha vakovlka se podobá vlčí tlapě, B - hrabavá noha vakokrta připomíná nohu krtka, C – přední noha koaly je upravena k přidržování se větví a poněkud připomíná prsty chameleona, D - vakoveverka páskovaná vytahuje larvy hmyzu ze dřeva prodlouženým 4. prstem podobně jako madagaskarská poloopice ksukol, E - noha klokana s redukcí prstů připomínající kopytníky (JK podle Veselovského, 1986).
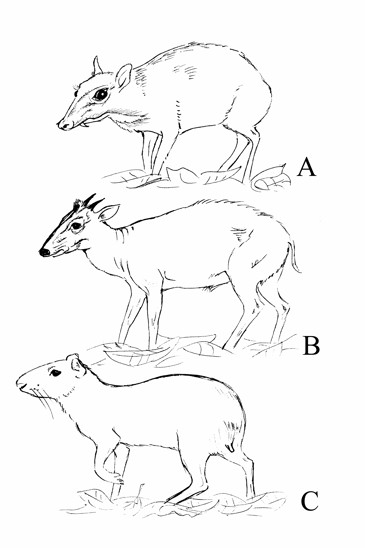
Obr. V podrostu tropických deštných lesů
paleotropické oblasti žijí kančilové (A), v Africe chocholatky (B) a neotropické
oblasti agutiové (C). V důsledku konvergentního vývoje vyhlížejí podobně, ač
kančilové patří do čeledě blízké jelenovitým, chocholatky mezi turovité a
agutiové jsou hlodavci (Obr. JK).

Obr. Letuchy (B) představují samostatný řád
(Dermoptera) a jsou spolu s tanami nejbližšími příbuznými primátů. Naproti tomu
poletuchy (A, C) patří do čeledi veverkovitých, tedy mezi hlodavce. Vakoveverka
(D) je australský vačnatec. Schopnost pasivního letu je u těchto vzájemně
nepříbuzných savců výsledkem konvergentního vývoje (Obr. JK).
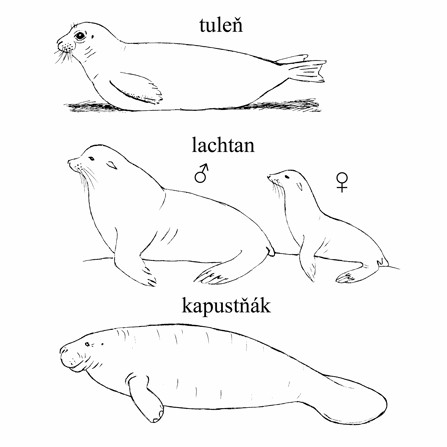
Obr. Tuleni a lachtani patří do řádu šelem,
kdežto kapustňák do řádu sirén. Příklad konvergentního vývoje (Obr. JK).
Jako
disparitu označujeme různorodost
(heterogenitu) tělních plánů popř. způsobů života uvnitř dané skupiny. Velkou
morfologickou disparitou se vyznačují např. měkkýši, malou např. hlístice.
S adaptací na mnohdy velmi prudce a drasticky se měnící podmínky se obecně lépe
vyrovnávají nepříliš specializovaní živočichové –
generalisté. Naopak úzce
specializované formy bývají málo adaptabilní a při změnách podmínek snadněji
zanikají.
Na tomto místě vhodné stručně
shrnout
základní geochronologické
pojmy (epochy, periody, éry).
Fylogenetika je věda
zabývající se rekonstrukcí historického vývoje organismů, studiem stupně jejich
vzájemné příbuznosti a vymezením taxonů vyšších než druh na základě sdílených
homologních znaků. Rozlišují se 3
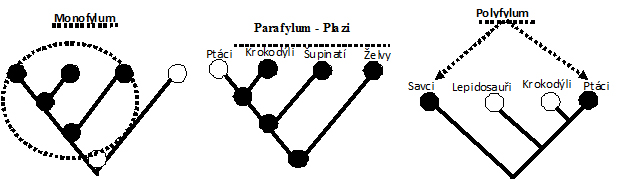
druhy taxonů s ohledem na jejich přirozenost - taxony monofyletické,
parafyletické a polyfyletické.
Taxon
je určitá skupina vzájemně příbuzných organizmů, které může být přiřazena určitá
taxonomická čili systematická jednotka neboli kategorie (druh, rod, čeleď apod.)
a která tvoří konkrétní náplň této jednotky. Jinými slovy je to pojmenovaná
taxonomická jednotka na jakékoli úrovni. Taxonem je tedy např. želvy bahenní,
želvy, plazi, strunatci.
Monofyletický taxon
(monofylum)
zahrnuje společného předka a všechny jeho potomky. Z fylogenetického pohledu se
jedná o jedinou přirozenou a správnou formu taxonu.
Parafyletický taxon
(parafylum)
zahrnuje společného předka, ale ne všechny jeho potomky (byl předkem i taxonu
jiného). Příkladem parafyletického taxonu jsou např. plazi, neboť v tradičním
pojetí nezahrnují ptáky, kteří s nimi sdílejí společného předka (ptáci a
krokodýli navíc tvoří společně taxon nazývaný Archosauria).
Polyfyletický taxon
(polyfylum)
je sběrná skupina založená na zdánlivé, ale nezávisle vzniklé podobnosti.
Příkladem polyfyletického taxonu mohou být
homoiotherma (živočichové se stálou
teplotou těla, tj. savci a ptáci) nebo
Pachydermata (tzv. tlustokožci, čirý taxonomický nesmysl). Existence taxonů
spojujících ptáky a savce, tedy taxonomických jednotek bez společného
nejbližšího předka, nemá opodstatnění.
Stejně nesmyslné je spojovat slony, nosorožce a hrochy jen proto, že mají silnou
kůži. Polyfyletické taxony vznikaly v minulosti, kdy zoologové neměli
k dispozici pokročilejší diagnostické metody.
Zatímco tradiční
evoluční taxonomie parafyletické
taxony uznává, moderní a dnes převažující
kladistická neboli
fylogenetická taxonomie je odmítá.
Polyfyletické taxony neakceptuje ani jedna z nich.
Podobnosti mezi znaky a taxony
Homologie
je podobnost
částí těla nebo orgánů různých organismů evolučně odvozená z odpovídajících
částí těla nebo orgánů společného předka (např. ruka člověka, hrudní končetina
koně, hrudní ploutev delfína, křídlo ptáka nebo netopýra, což vše jsou
modifikace odvozené ze společného embryonálního základu). Homologie můžeme
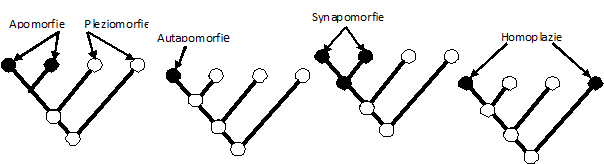
rozdělit na podobnost pleziomorfní a apomorfní.

Obr. Křídlo netopýra je orgán homologický např.
s lidskou rukou nebo křídlem ptáka, neboť jde o modifikace odvozené ze
společného evolučního základu (hrudní končetiny primitivních tetrapodů).
Současně jde o projev konvergence s ptačím křídlem, neboť vzniklo v procesu
adaptace na obdobnou ekologickou niku (Foto ZL).
Pleziomorfie
je shoda
v původním (primitivnějším) stavu znaku (např. pětiprsté končetiny u tetrapodů).
Apomorfie
je přítomnost
evoluční novinky, tj. je stavem odvozeným (např. křídla ptáků). U apomorfií
můžeme ještě rozlišovat autapomorfie (jedinečný výskyt apomorfie, např.
přeměna předního páru křídel u brouků na krovky je znak pro brouky autapomorfní)
a synapomorfie, týkající se společných, sdílených apomorfií (sdílení
evoluční novinky, např. přítomnost křídel u brouků je synapomorfie, tj. sdílená
apomorfie s ostatním křídlatým hmyzem, poněvadž bezkřídlost se považuje za
původní stav, pleziomorfii).
Další příklady:
je-li stavem znaku přítomnost či absence zubů, pak bezzubost ptáků či želv je
apomorfií (jejich předkové zuby měli). Přítomnost zubů u ostatních amniot (plazi
v tradičním pojetí a savci) je pleziomorfií, neboť jejich předkové
(obojživelníci) rovněž měli zuby.
Analogie,
homoplazie, konvergence
- neboli nehomologická podobnost je
povrchní podobnost funkčního charakteru u fylogeneticky nepříbuzných skupin
(např. křídla hmyzu a ptáků, která se vyvinula z rozdílných výchozích struktur).
Je to fenotypová podobnost znaků u taxonů, které
nemají blízkého společného předka a
tedy ani společný evoluční základ/původ (ryby a delfíni). Její existence
komplikuje práci taxonomů a je příčinou, proč dochází k neustálým změnám v
biologické systematice, neboť starší systémy byly mnohdy založeny na pouhé
podobnosti, nikoli skutečné
příbuznosti organismů.
Reverze
- vývoj znaku ke stavu připomínajícímu původní stav u vzdálených předků
(druhotná ztráta křídel u hmyzu, např. vší nebo blech, připomínající původní
bezkřídlé primitivní formy).
Typy dat pro konstrukci fylogenetického stromu
Data používaná pro
konstrukci fylogenetických stromů mohou mít buď
formu vlastních znaků (např.
přítomnost pětiprstých končetin nebo křídel, druh nukleotidu na konkrétní pozici
v genové sekvenci) nebo se může jednat o data
ve formě vzdáleností (distancí) či
podobností, což jsou vypočítané
číselné údaje udávající např. procentuální rozdílnost studovaných taxonů ve
sledovaných znacích. Znaky mohou být jak kvalitativní tak kvantitativní a mohou
být převedeny na distance či podobnosti, opačný směr (z distancí na znaky) není
možný (např. distanci 13,28 nelze převést na přítomnost či absenci pětiprstých
končetin u tetrapodů či Stiedova tělíska u kokcidií). Významnými podmínkami pro
volbu sledovaných znaků
(morfologických, biochemických, imunologických, cytogenetických nebo molekulárně
genetických), jsou jejich vzájemná
nezávislost (např. přeměna pětiprstých končetin v křídla ptáků nijak
nesouvisí se ztrátou zubů a přítomností zobáku, jedná se tedy o dva nezávislé
sledovatelné znaky) a pro jednotlivá znaková data také jejich
homologie. Z populárních a často
používaných metod analýzy nukleových kyselin poskytuje pouze DNA-DNA hybridizace
data výhradně ve formě vzdáleností či podobností. U znaků ještě rozlišujeme
stav znaku. Pro snazší pochopení si
lze u molekulárně genetických, fylogenetických studií představit jako
znak nukleotid v sekvenci
konkrétního studovaného genu na určité pozici (např. 550), jako
stav znaku je konkrétní nukleotid
(A, T, C nebo G).
Grafickým
znázorněním výsledků fylogenetické studie je fylogenetický strom. Tento strom
může být zakořeněný (kořen představuje společného předka všech
studovaných taxonů) nebo nezakořeněný (nejstarší společný předek není
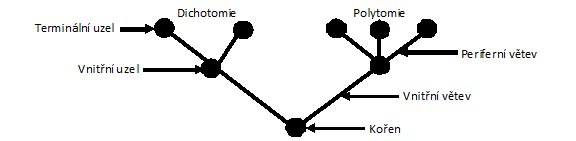
identifikován). Fylogenetický strom se skládá z větví a uzlů, kdy
terminální uzly představují recentní taxony a vnitřní uzly
představují hypotetické předky. U kořene leží
bazální větve vznikající při prvních
divergencích, zatímco terminální větve se štěpí v
korunové skupiny s převahou
odvozených znaků. V protikladu k nim označujeme bazálněji ležící větve s
převahou pleziomorfních znaků jako
kmenové skupiny. Nejblíže příbuzné skupiny sdílející společného předka se
nazývají sesterské. Jestliže vnitřní
uzel spojuje tři větve, jedná se o dichotomické větvení (dichotomii),
kdy se jedna původní
větev ve vnitřním uzlu dělí na dvě následné. Spojuje-li vnitřní uzel více větví,
jedná se o polytomické větvení (polytomii). Polytomie by se v dobře
vyřešených fylogenetických stromech prakticky neměly vyskytovat. Je totiž velmi
nepravděpodobné, že by se jedna evoluční linie rozpadala v jednom okamžiku na
více než dvě nové evoluční linie. Polytomie tak nejčastěji vznikají neschopností
zachytit rychle po sobě následující dichotomická větvení.
Ke znázornění
různých aspektů evoluce lze použít různé druhy fylogenetických stromů:
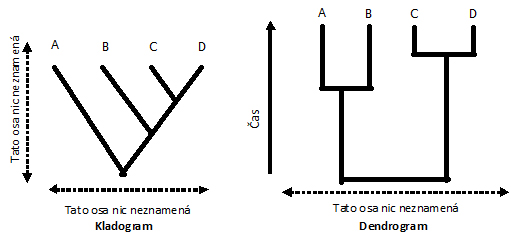
Kladogram
(z řec. klados = větev) je
základní typ stromu, který popisuje příslušnou hypotézu o příbuznosti, aniž by
řešil, kdy došlo k štěpení jednotlivých linií nebo do jaké míry se jednotlivé
evoluční linie liší.
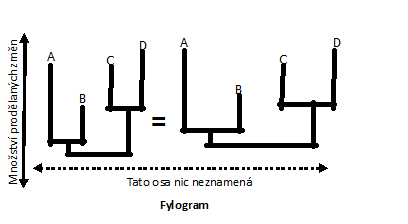
Fylogram
(aditivní strom)
podává dodatečné (aditivní) informace ve formě délky větví. Délka větve tak
odpovídá například množství prodělaných evolučních změn. Rychlost mutací a
vzniku evolučních novinek totiž není konstantní pro všechny. Záleží na
prostředí, délce trvání generačního intervalu a mnoha dalších faktorech.
Dendrogram
(ultrametrický
strom) představuje další formu grafického znázornění (je jedním z aditivních
stromů), kdy koncové uzly představující recentní druhy jsou ve stejné rovině
(představující současnost), stejně vzdálené od společného předka. Dendrogram ( z
řec. dendron = strom) popisuje průběh evolučních změn v čase, z polohy uzlů lze
odečíst kdy došlo ke štěpení jednotlivých evolučních linií.
Nejjednodušší
metodou
využívanou ke konstrukci fylogenetických stromů je metoda
parsimonie. Jedná se o hledání
nejjednoduššího řešení a preferováním jednodušších hypotéz před složitějšími.
Tyto výukové materiály byly spolufinancovány Evropským sociálním fondem a státním rozpočtem ČR.